
Wie verändert sich die Biodiversität in Zeit und Raum und was sind die treibenden Kräfte dahinter? Eine große internationale Gruppe (Alroy et al. 2008) hat die traditionelle Ansicht des exponentiellen Wachstums der marinen Biodiversität in den letzten 200 Millionen Jahren (siehe Abbildung) revidiert. Die Erkenntnis basiert auf einer großen Datenbank mit Informationen über den gesamten Fossilbericht.
Die Paleobiology Database erlaubt es, moderne Methoden der Probenstandardisierung anzuwenden, um biologisch aussagekräftige Messwerte der vergangenen Dynamik der Biodiversität zu erhalten. Diese Datenbank wird kontinuierlich von einem internationalen Team von Wissenschaftlern, nicht zuletzt von Mitarbeitern des Geozentrums Nordbayern, weiterentwickelt. Unsere Gruppe nutzt die Datenbank um die Dynamik der Biodiversität in Raum und Zeit zu untersuchen. Eine wichtige Fragestellung betrifft das Zusammenspiel von biotischen und abiotischen Faktoren, die die Diversifikation des Lebens kontrollieren.
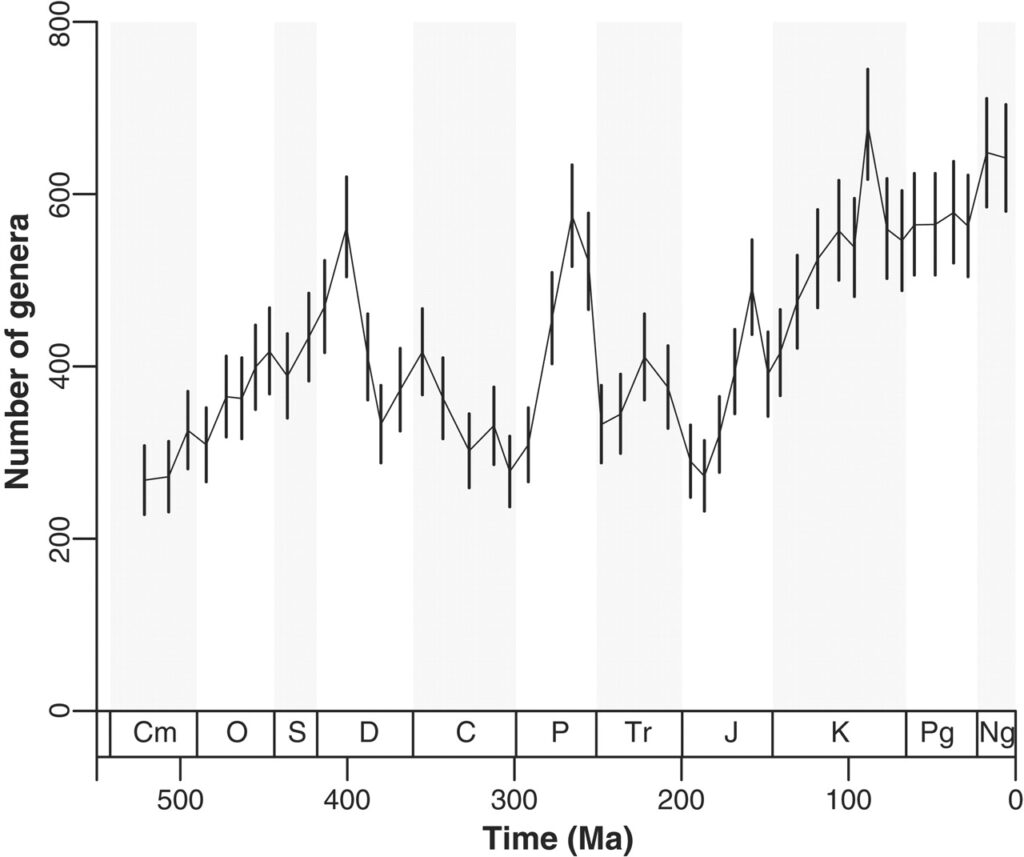
Abbildung: Artenvielfalt auf Gattungsebene sowohl bei heute lebenden als auch bei ausgestorbenen wirbellosen Meerestieren (Metazoen ohne Tetrapoden) während des Phanerozoikums, basierend auf einer stichprobenstandardisierten Analyse der Paleobiology Database.
Literatur
Alroy J, et al. (2008) Phanerozoic trends in the diversity of marine invertebrates. Science 321:97-100.
Wolfgang Kiessling, Carl Simpson
Obwohl die Symbiose zwischen Algen und Korallen oft als ökologischer Vorteil für beide Seiten angesehen wird, ist der evolutionäre Erfolg von Steinkorallen nicht unbedingt durch die Photosymbiose bedingt. Die heute etwa gleiche Anzahl von zooxanthellaten und azooxanthellaten Korallenarten (beide haben etwa 750 existierende Arten) deutet eher auf einen makroevolutionären Kompromiss hin.
In einem von der DFG geförderten Projekt kombinieren wir zeitkalibrierte molekulare und fossile Daten, um eine bestmögliche Schätzung der Diversifizierungsraten verschiedener ökologischer Lebensweisen von Steinkorallen im Laufe der Zeit abzuleiten.
Für Riffkorallen konnten wir bereits zeigen, dass die Ergebnisse für fossile und molekulare Daten ähnlich sind, sodass die Daten kombiniert werden können, um unverzerrte Raten abzuleiten (Simpson et al. 2011). Das gleiche Instrumentarium wird nun auf azooxanthellate Korallen angewendet, für die es viel weniger Fossilienfunde gibt. (übersetzt)

Abbildung: Acropora, die heute am weitesten verbreitete zooxanthellate Riffkoralle (Rotes Meer, Ägypten).
Wolfgang Kiessling, Michael Joachimski, Axel Munnecke
„Einer der größten Vorzüge der Paläontologie ist ihre Fähigkeit, das Ausmaß und den Zeitpunkt früherer Aussterbeereignisse direkt zu bewerten, sofern die Verzerrungen der Fossilienfunde berücksichtigt werden. Während zeitkalibrierte molekulare Daten zur Bewertung der Diversifizierungsraten herangezogen werden können, liefern nur Fossilienfunde Informationen über frühere Aussterbeereignisse. Die mit Abstand quantitativsten Daten zu Aussterbeereignissen stammen aus den marinen Fossilienfunden in Schelfhabitaten. Dies ist genau der Lebensraum, für den die aktuellen Aussterberaten am schwierigsten zu bewerten sind und für den daher nur unzureichende Daten vorliegen. Eines der wichtigsten Muster, das sich aus den Fossilienfunden ergibt, ist, dass alte Massenaussterben in der Regel mit Zeiten der globalen Erwärmung und nicht, wie bisher angenommen, mit Zeiten der Abkühlung zusammenhängen (siehe Abbildung).
Mehrere Projekte am GeoZentrum befassen sich mit den Mustern, Ursachen und evolutionären Folgen mariner Massenaussterben. Die Muster werden anhand detaillierter Feldarbeiten in Kombination mit geochemischen Analysen und Datenbankanalysen bewertet. Unsere Schwerpunkte sind das Massensterben am Ende des Ordoviziums (Delabroye et al. 2011), die Bioereignisse im Silur (Munnecke et al. 2012), das Massensterben am Ende des Perm und zu Beginn der Trias (Joachimski et al. 2012), das Massensterben am Ende der Trias (Kiessling et al. 2007, 2009) und das Massensterben am Ende der Kreidezeit (Aberhan et al. 2007; Schulte et al. 2010).“ (übersetzt)
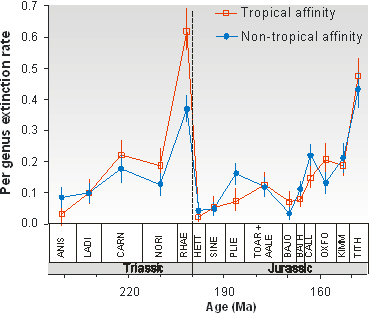
Abbildung: Tropical genera were hit significantly harder during the end-Triassic mass extinction that non-tropical genera. This suggests that global warming was an important trigger of the extinctions (from Kiessling and Aberhan 2007).
Literatur
Aberhan M, Weidemeyer S, Kiessling W, Scasso R, & Medina FA (2007) Faunal evidence for reduced productivity and uncoordinated recovery in Southern Hemisphere Cretaceous/Paleogene boundary sections. Geology 35:227-230.
Delabroye A, et al. (2011) Phytoplankton dynamics across the Ordovician/Silurian boundary at low palaeolatitudes: Correlations with carbon isotopic and glacial events. Palaeogeography, Palaeoclimatology, Palaeoecology 312:79-97.
Joachimski MM, et al. (2012) Climate warming in the latest Permian and the Permian-Triassic mass extinction. Geology 40:195-198.
Kiessling W & Aberhan M (2007) Environmental determinants of marine benthic biodiversity dynamics through Triassic-Jurassic times. Paleobiology 33:414-434.
Kiessling W, Aberhan M, Brenneis B, & Wagner PJ (2007) Extinction trajectories of benthic organisms across the Triassic-Jurassic boundary. Palaeogeography, Palaeoclimatology, Palaeoecology 244:201-222.
Kiessling W, Roniewicz E, Villier L, Leonide P, & Struck U (2009) An early Hettangian coral reef in southern France: Implications for the end-Triassic reef crisis. Palaios 24:657-671.
Munnecke A, Delabroye A, Servais T, Vandenbroucke TRA, & Vecoli M (2012) Systematic occurrences of malformed (teratological) acritarchs in the run-up of Early Palaeozoic δ13C isotope excursions. Palaeogeography, Palaeoclimatology, Palaeoecology 367-368:137-146.
Schulte P, et al. (2010) The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary. Science 327:1214-1218.
Wolfgang Kiessling, Christoph Heubeck, Michael Steiner, Zhu Maoyan, Li Guoxiang, Na Lin
„The Cambrian period marks the appearance of modern animal phyla in the fossil record. It also marks the origin of organisms with skeletons, the invention of complex food webs, which are in many similar to modern ones, and an increase of global biodiversity. Because of the geologically sudden appearance of these events these changes are commonly referred to as the Cambrian Explosion. Geological, ecological and genetic factors are discussed as causes of the Cambrian Explosion. Embedded in an international Research Group (Opens external link in new windowFOR 736), our team in Erlangen explores global and regional biodiversity dynamics during Ediacaran-Cambrian times. Biodiversity dynamics are analysed to (1) uncover the timing of major changes on different continents and (2) to assess if the Cambrian bioradiation was triggered mostly by an increase of within-community diversity (alpha diversity) or rather by an increasing difference among communities (beta diversity).“Die Kambrium-Periode markiert das Auftreten moderner Tierstämme in den Fossilienfunden. Sie markiert auch den Ursprung von Organismen mit Skeletten, die Entstehung komplexer Nahrungsnetze, die in vielerlei Hinsicht den heutigen ähneln, und eine Zunahme der globalen Artenvielfalt. Aufgrund des geologisch plötzlichen Auftretens dieser Ereignisse werden diese Veränderungen gemeinhin als die Kambrium-Explosion bezeichnet. Als Ursachen für die Kambrium-Explosion werden geologische, ökologische und genetische Faktoren diskutiert. Eingebettet in eine internationale Forschungsgruppe (Öffnet externen Link in neuem FensterFOR 736) untersucht unser Team in Erlangen die globale und regionale Biodiversitätsdynamik während des Ediacarium-Kambrium. Die Biodiversitätsdynamik wird analysiert, um (1) den Zeitpunkt wichtiger Veränderungen auf verschiedenen Kontinenten aufzudecken und (2) zu beurteilen, ob die kambrische Bioradiation hauptsächlich durch eine Zunahme der Vielfalt innerhalb der Gemeinschaften (Alpha-Diversität) oder eher durch eine zunehmende Differenzierung zwischen den Gemeinschaften (Beta-Diversität) ausgelöst wurde.“ (übersetzt)

Abbildung: Kegelkarst in präkambrischen und kambrischen Sedimentgesteinen Südchinas (Provinz Guizhou)